Recentemente tem surgido na internet textos com o seguinte “motor”: “dietas de restrição de açúcar curam o câncer”, ou, mais tecnicamente, “dietas cetogênicas curam o câncer”.
Lembrando o infeliz caso da fosfoetanolamina, é de se perguntar sempre, mesmo que a notícia seja extremamente séria e oriunda de grande (e caro) esforço de pesquisa médica: Qual câncer?
Lembrando o infeliz caso da fosfoetanolamina, é de se perguntar sempre, mesmo que a notícia seja extremamente séria e oriunda de grande (e caro) esforço de pesquisa médica: Qual câncer?
No texto a seguir, não tentaremos de forma alguma afirmar que dietas com redução da ingestão de carboidratos não colaborem para tratamentos de câncer, o que ficará bem destacado inclusive em artigo de divulgação da área média que iremos acrescentar.
COmo seguidamente divulgo, e me informei sobre tal ainda no início dos anos 90, a ingestão elevada de açúcar, ainda mais se somada a grande ingestão de café (o café muito açucarado) é suspeito há anos pelos médicos de ser um dos causadores do chamado “câncer da cabeça do pâncreas”.
Notemos: açúcar elevando a taxa de glicose no sangue, mais alta produção de insulina no pâncreas para enfrentar esta glicose, uma fonte de diversas substâncias a serem metabolizadas (o café torrefado), atuando também como um meio que causa o constante consumo ao longo do dia, levando-nos a perceber que pode haver um nexo causal na síndrome neste caso.
Mas aqui não trataremos da causa que leva ao surgimento de células cancerosas, que incluem até a ingestão elevada de gorduras e toda a bioquímica da decomposição do glicerol (sempre presente em gorduras) pelo aquecimento, ou mesmo, as diversas associações entre obesidade e câncer que tem sido feitas.
Trataremos da relação causa efeito de ao se cortar um ingrediente de nossa nutrição, isso conduzir às células cancerosas a morrerem, pois disso que se trata “curar câncer”.
COmo seguidamente divulgo, e me informei sobre tal ainda no início dos anos 90, a ingestão elevada de açúcar, ainda mais se somada a grande ingestão de café (o café muito açucarado) é suspeito há anos pelos médicos de ser um dos causadores do chamado “câncer da cabeça do pâncreas”.
Notemos: açúcar elevando a taxa de glicose no sangue, mais alta produção de insulina no pâncreas para enfrentar esta glicose, uma fonte de diversas substâncias a serem metabolizadas (o café torrefado), atuando também como um meio que causa o constante consumo ao longo do dia, levando-nos a perceber que pode haver um nexo causal na síndrome neste caso.
Mas aqui não trataremos da causa que leva ao surgimento de células cancerosas, que incluem até a ingestão elevada de gorduras e toda a bioquímica da decomposição do glicerol (sempre presente em gorduras) pelo aquecimento, ou mesmo, as diversas associações entre obesidade e câncer que tem sido feitas.
Trataremos da relação causa efeito de ao se cortar um ingrediente de nossa nutrição, isso conduzir às células cancerosas a morrerem, pois disso que se trata “curar câncer”.
Abordagem pelos aminoácidos como fonte de nutrição
Todo bom praticante de musculação, um aficionado como eu, e ainda mais os fisiculturistas, sabem que se não contrabalançar a catabolia pós-treino com carboidratos, o corpo requererá aminoácidos - imprescindíveis para a construção de músculos - em energia, e nisso, se terá desperdícios da nutrição e obviamente, menos ganhos musculares.
Qualquer texto de divulgação sobre a metabolização dos aminoácidos os mostra como fonte de energia pelo organismo em caso de necessidade (no caso de nós, primatas).
Qualquer texto de divulgação sobre a metabolização dos aminoácidos os mostra como fonte de energia pelo organismo em caso de necessidade (no caso de nós, primatas).
“Peculiar para estas biomoléculas, é a presença do grupo amino. Portanto, cada via degradativa passa por um passo-chave, no qual o grupo a-amino é separado do esqueleto carbônico e desviado para uma via especializada para o metabolismo do grupamento amino. Os alfa-cetoácidos assim formados, em geral, encaminham-se para o Ciclo do Ácido Cítrico, podendo sofrer oxidação até CO2 e H2O, ou ainda, seus esqueletos carbônicos podem fornecer unidades de três e quatro átomos de carbono para a conversão em glicose.” - Degradação de Aminoácidos - www.lbqp.unb.br
Reforcemos:
What is there in the cell which codes for the carbohydrate sequence in the cells? - Sai Janani Ganesan - www.quora.com
Destaque-se:
“We don't necessarily need carbohydrates in our diet. We can convert some amino acids/organic molecules into carbohydrates (CH) via Gluconeogenesis”
( Nós não precisamos necessariamente de carboidratos em nossa dieta. Podemos converter alguns aminoácidos / moléculas orgânicas em carboidratos (CH) via gliconeogênese. )
Em suma: aminoácidos são transformados em glicose.
Uma observação biológica dos cordados: nossos cérebros “funcionam” apenas com glicose, de onde é uma necessidade manter um estoque de glicose no sangue e em outras formas de reserva para abastecer o cérebro, sem o que nada pode funcionar. Por isso que corredores de fundo, nadadores de longas distâncias e triatletas a partir de determinado momento queimam gordura - na verdade, qualquer um o faz após aproximadamente uns 30 minutos mesmo caminhando tranquilamente numa esteira - mas a cada tanto tempo, necessitam de uma certa ingestão de algo açucarado, e como profissional da química, devo alertar que mais que “açucarado”, com glicose presente.
Todos que já tiveram alguma tremedeira, suor frio, dificuldade de raciocínio e até visão em túnel sabem dos efeitos da falta de glicose. Desmaios em plena academia veem de exemplo mais forte.
Logo, não ingerir carboidratos não elimina a presença de glicose no nosso corpo.
Somando outro ponto, nos anos 60 e 70, fisiculturistas de ponta nos EUA recomendavam dietas cetogênicas para preparação para competições, dado seu forte efeito de redução da gordura corporal, a própria notória e bem ilustrativa “enciclopédia” de Arnold Schwarzenegger faz recomendações sobre o tema. O que é sempre alertado são os “efeitos tóxicos” deste tipo de dieta.
Uma rápida pesquisa pela internet sobre o tema já mostra assustadoras afirmações:
“Apesar de realmente proporcionar um emagrecimento rápido, a dieta cetogênica favorece uma série de problemas de saúde como câncer, hipoglicemia, colesterol alto, fadiga, dores de cabeça, entre outros.” - Bruna Stuppiello; Dieta cetogênica: alimentos permitidos, receitas, vantagens e desvantagens - Método proporciona emagrecimento rápido, mas é preciso cuidado pois pode favorecer problemas de saúde - www.minhavida.com.br
Uma observação biológica dos cordados: nossos cérebros “funcionam” apenas com glicose, de onde é uma necessidade manter um estoque de glicose no sangue e em outras formas de reserva para abastecer o cérebro, sem o que nada pode funcionar. Por isso que corredores de fundo, nadadores de longas distâncias e triatletas a partir de determinado momento queimam gordura - na verdade, qualquer um o faz após aproximadamente uns 30 minutos mesmo caminhando tranquilamente numa esteira - mas a cada tanto tempo, necessitam de uma certa ingestão de algo açucarado, e como profissional da química, devo alertar que mais que “açucarado”, com glicose presente.
Todos que já tiveram alguma tremedeira, suor frio, dificuldade de raciocínio e até visão em túnel sabem dos efeitos da falta de glicose. Desmaios em plena academia veem de exemplo mais forte.
Logo, não ingerir carboidratos não elimina a presença de glicose no nosso corpo.
Somando outro ponto, nos anos 60 e 70, fisiculturistas de ponta nos EUA recomendavam dietas cetogênicas para preparação para competições, dado seu forte efeito de redução da gordura corporal, a própria notória e bem ilustrativa “enciclopédia” de Arnold Schwarzenegger faz recomendações sobre o tema. O que é sempre alertado são os “efeitos tóxicos” deste tipo de dieta.
Uma rápida pesquisa pela internet sobre o tema já mostra assustadoras afirmações:
“Apesar de realmente proporcionar um emagrecimento rápido, a dieta cetogênica favorece uma série de problemas de saúde como câncer, hipoglicemia, colesterol alto, fadiga, dores de cabeça, entre outros.” - Bruna Stuppiello; Dieta cetogênica: alimentos permitidos, receitas, vantagens e desvantagens - Método proporciona emagrecimento rápido, mas é preciso cuidado pois pode favorecer problemas de saúde - www.minhavida.com.br
Uma breve pausa para insistir no nosso argumento
Na mesma fonte acima, encontramos:
“Dieta cetogênica e câncer
Na mesma fonte acima, encontramos:
“Dieta cetogênica e câncer
Muitas pessoas acreditam que a dieta cetogênica pode ser uma aliada do tratamento do câncer. Isso acontece, principalmente, devido ao fato de que enquanto células normais conseguem entrar em cetose - ou seja, usar os ácidos graxos no lugar da glicose como fonte de energia, quando não há consumo de carboidratos - as células cancerígenas são incapazes de fazer o mesmo.
No entanto, por mais que o corpo não esteja ingerindo glicose via alimentação, ainda há produção de glicose pelas células e aparentemente as células cancerígenas conseguem viver e se reproduzir nessas condições.
Em um estudo conduzido em 2012 e publicado pela revista científica Nutrition, 10 pacientes com câncer avançado foram submetidos a uma dieta cetogênica por 28 dias. Depois do período, 4 pacientes estavam com a doença ainda progredindo, 5 estabilizaram a apenas um teve uma remissão parcial do tumor.
Por isso, ainda não dá para dizer com certeza se esse tipo de dieta realmente colabora para quem sofre com câncer, e mais estudos maiores precisam ser conduzidos para se chegar a uma conclusão sobre o tema.”
Abordagem pelo caráter “evolutivo” do que sejam as células cancerosas
Encontram-se facilmente autores que fazem uma abordagem “darwinista”, evolutiva, para o que seja o câncer, mais especificamente, o que sejam evolutivamente as células cancerígenas.
Não somos, assim como os demais seres pluricelulares, “um indivíduo”. Somos uma coletividade de células que formam o que chamamos, até por “hilemorfismo”, um indivíduo. Tente aplicar estas definições de individualidade a um cnidário de múltiplas genéticas, ou mesmo a um hidra com um brotamento e verá que a situação começará a ficar confusa. Mesmo uma simples planta de jardim já apresenta características que podem por em xeque a definição do que seja um indivíduo.
Mas voltemos a um primata de algum porte como eu.
Imaginemos que uma determinada mutação, ou mesmo a dificuldade de replicação do DNA causada pelo clássico benzopireno do carvão mineral provoque as modificações de comportamento que levam determinada célula do revestimento de meus intestinos a tornarem-se o que chamamos de célula cancerígena.
Ela começa a se comportar como um ser vivo extremamente primitivo, que apenas “deseja” se reproduzir e se nutrir do meio, no caso, abastecido pelo restante de meu corpo saudável.
Este meio provê os nutrientes que a abastece, reforcemos, como um leito lodoso de um rio abastece um grupo de amebas.
Mas voltemos a um primata de algum porte como eu.
Imaginemos que uma determinada mutação, ou mesmo a dificuldade de replicação do DNA causada pelo clássico benzopireno do carvão mineral provoque as modificações de comportamento que levam determinada célula do revestimento de meus intestinos a tornarem-se o que chamamos de célula cancerígena.
Ela começa a se comportar como um ser vivo extremamente primitivo, que apenas “deseja” se reproduzir e se nutrir do meio, no caso, abastecido pelo restante de meu corpo saudável.
Este meio provê os nutrientes que a abastece, reforcemos, como um leito lodoso de um rio abastece um grupo de amebas.
Logo, as terapias contra o câncer mais eficientes tratam de atacar tais células com radiação (radioterapia), envenená-las (quimioterapia), algumas drogas que as fazem ser detectadas pelo sistema imunológico, casos específicos e recentes em tratamentos com fármacos em que a própria reprodução e nutrição destas células é prejudicada e são levadas à morte, e até tratamentos ainda mais avançados e em pesquisa com uso de vírus e engenharia genética.
Mas notemos que se “fome” seja do que nutriente for resolvesse por si câncer, os pacientes de câncer de estômago, de intestino, de todo o sistema digestivo e até prejudicados por piores metabolizações no fígado, por exemplo, não teriam seus cânceres em avanço e muito menos as metástases que também seriam prejudicadas por este abalo de nutrição.
Aqui, o motivo principal é que tal como amebas, as células cancerígenas colocarão todo seu maquinário celular em trabalho para desesperadamente manter-se vivas, e chocante, elas evoluem e se adaptam adquirindo novas características, buscando novas nutrições possíveis na massa de ser vivo (eu, por exemplo) que as cercam.
Recomendamos:
Cancer Therapy: An Evolved Approach -
Mas notemos que se “fome” seja do que nutriente for resolvesse por si câncer, os pacientes de câncer de estômago, de intestino, de todo o sistema digestivo e até prejudicados por piores metabolizações no fígado, por exemplo, não teriam seus cânceres em avanço e muito menos as metástases que também seriam prejudicadas por este abalo de nutrição.
Aqui, o motivo principal é que tal como amebas, as células cancerígenas colocarão todo seu maquinário celular em trabalho para desesperadamente manter-se vivas, e chocante, elas evoluem e se adaptam adquirindo novas características, buscando novas nutrições possíveis na massa de ser vivo (eu, por exemplo) que as cercam.
Recomendamos:
Cancer Therapy: An Evolved Approach -
Tumours are subject to the same rules of natural selection as any other living thing. Clinicians are now putting that knowledge to use - Cassandra Willyard, Nature; April 13, 2016 - www.scientificamerican.com
( Os tumores estão sujeitos às mesmas regras de seleção natural que qualquer outra coisa viva. Os médicos agora estão colocando esse conhecimento em uso )
Evolution Research Could Revolutionize Cancer Therapy
Evolutionary studies indicate that the genetic changes enabling a cancer to develop arise shockingly early within the primary tumor. This discovery points to a promising new approach to therapy - Jeffrey P. Townsend - www.scientificamerican.com
( Estudos evolutivos indicam que as mudanças genéticas que permitem o desenvolvimento de um câncer surgem de forma chocante no início do tumor primário. Esta descoberta aponta para uma nova abordagem promissora para a terapia )
O argumento do câncer em animais carnívoros
Agora pensemos: se a ausência de glicose por si pudesse curar câncer, animais carnívoros por excelência se curariam naturalmente de seus tumores.
Qualquer pessoa que convive com cães e gatos (e gatos são um exemplo mais marcante de carnivorismo) sabem que eles adquirem cânceres. Entre os cães pit bull, é bem conhecido seu câncer de pele, normalmente propulsionado pelo ultravioleta da luz solar, o que nos leva às perguntas “Qual o motor do câncer mesmo?” e “Qual tipo de câncer?”, e não a causas únicas que não podem ser afirmadas, característica únicas do câncer que são muito mais amplas que apenas o uso do termo, constância de comportamento que como vimos não se sustentam, e lógico, abordagens para tratamento (se ele houver!) que jamais passarão por uma panacéia.
Qualquer pessoa que convive com cães e gatos (e gatos são um exemplo mais marcante de carnivorismo) sabem que eles adquirem cânceres. Entre os cães pit bull, é bem conhecido seu câncer de pele, normalmente propulsionado pelo ultravioleta da luz solar, o que nos leva às perguntas “Qual o motor do câncer mesmo?” e “Qual tipo de câncer?”, e não a causas únicas que não podem ser afirmadas, característica únicas do câncer que são muito mais amplas que apenas o uso do termo, constância de comportamento que como vimos não se sustentam, e lógico, abordagens para tratamento (se ele houver!) que jamais passarão por uma panacéia.
Tipos de câncer mais frequentes em gatos - meusanimais.com.br
Bom, primatas (eu) não são cães e gatos, evidentemente, mas a questão parece bem clara, até porque, como mostramos “evolutivamente” acima, as células cancerígenas não são as células de um primata propriamente dita, assim como não são no cão células caninas ou num gato células felinas. Exatamente deixarem de ser as células “comuns” destes seres vivos é que é o argumento por nosso ponto que cortar um nutriente - ou um espectro de nutrientes - não pode matá-las banalmente.
Uma segunda opinião...
A seguir, acrescentamos a tradução de um texto de divulgação bem didático e abrangente sobre o tema, que acredito, eliminará diversas mistificações sobre o tema, e espero, reforce o que apresentei.
Sugar and cancer – what you need to know - Emma Smith - May 15, 2017 - scienceblog.cancerresearchuk.org
Açúcar e câncer - o que você precisa saber
Este post foi atualizado em 9 de novembro de 2018 com as informações mais recentes sobre como o governo está lidando com o açúcar.
Há muitas informações e conselhos confusos ao redor do açúcar.
Ele se tornou o vilão da nossa dieta, mas onde está o consenso entre como o açúcar e o câncer estão ligados?
Isso causa câncer? O açúcar alimenta as células cancerígenas, fazendo-as crescer de forma mais agressiva? E como o açúcar que consumimos através da comida e bebida afeta nossa saúde e o que pode ser feito a respeito disso?
Neste post, estaremos analisando muito o açúcar.
Vamos nos concentrar especificamente no açúcar e no câncer, quebrando alguns mitos e cobrindo o que os pesquisadores estão estudando na esperança de encontrar novas maneiras de tratar as pessoas com câncer.
E nós vamos cobrir porque a quantidade de açúcar em nossas dietas é motivo de preocupação. Uma dieta rica em açúcar pode ser uma má notícia quando se trata de risco de câncer, mas não pelas razões que muitas vezes aparecem nas manchetes.
Mas primeiro o básico, para o que nossos corpos precisam de açúcar e de onde vem em nossa dieta.
Glicose - o combustível da vida
Pesquise açúcar e câncer na internet e não demore para encontrar alertas alarmantes de que o açúcar é a "morte branca" e a "comida favorita do câncer".
Mas essa ideia de que o açúcar é responsável por dar o pontapé inicial ou alimentar o crescimento de um câncer é uma simplificação excessiva de alguma biologia complicada. Vamos começar com o que o açúcar realmente é.
Açúcar vem em muitas formas diferentes. A forma mais simples é como uma única molécula, como glicose e frutose. Essas moléculas de açúcares simples também podem se unir, em pares ou como cadeias mais longas de moléculas. Todas essas combinações de moléculas são carboidratos e são a principal fonte de energia do nosso corpo.
A forma de açúcar com a qual a maioria de nós estará familiarizada é o açúcar de mesa, que é um açúcar simples que se dissolve na água e dá um sabor doce às coisas. Seu nome próprio é sacarose, e é composto de cristais de glicose e frutose. O açúcar de mesa é refinado, o que significa que foi processado para extraí-lo de uma fonte natural (geralmente beterraba sacarina). Os alimentos não transformados também podem ter alto teor de açúcares simples, por exemplo, o mel (também feito principalmente de glicose e frutose) é quase puro açúcar.
À medida que as cadeias de açúcar aumentam, elas perdem o sabor adocicado e não se dissolvem mais na água. Essas cadeias são chamadas de polissacarídeos e formam um grande componente de alimentos ricos em amido. Alimentos ricos em amido, como arroz, pão, macarrão e legumes, como batatas, podem não ter sabor doce, mas também são ricos em carboidratos.
Açúcar, de alguma forma, é em muitas coisas que comemos. E isso é bom, porque nossos corpos dependem muito dele para funcionar.
Quase todas as partes do nosso corpo são feitas de células vivas. E são essas células que nos ajudam a ver, respirar, sentir, pensar e muito mais.
Embora seus empregos no corpo possam diferir, uma coisa que todas essas células têm em comum é que elas precisam de energia para sobreviver e desempenhar suas funções.
As células precisam, de alguma forma, transformar nutrientes em nossa dieta em uma forma de energia que eles possam usar, chamada ATP. Levaria muito tempo para explicar isso (se você estiver interessado, talvez queira ler mais), mas de maneira simplista, o processo começa com a glicose.
A glicose é o combustível básico que alimenta todas as nossas células. Se comermos ou bebermos coisas com alto teor de glicose, como bebidas com gás, a glicose é absorvida diretamente em nosso sangue, pronta para uso por nossas células. Se um alimento rico em amido, como macarrão, estiver no cardápio, as enzimas em nossa saliva e sucos digestivos quebram e convertem em glicose. E se por algum motivo não houver carboidrato em nossa dieta, as células podem transformar gordura e proteína em glicose como último recurso, porque precisam de glicose para sobreviver.
É aqui que o açúcar e o câncer começam a colidir, porque o câncer é uma doença das células.
Açúcar e Câncer
As células cancerosas geralmente crescem rapidamente, multiplicando-se rapidamente, o que requer muita energia. Isso significa que eles precisam de muita glicose. As células cancerosas também precisam de muitos outros nutrientes, como aminoácidos e gorduras; não é só o açúcar que eles desejam.
É aí que nasceu o mito de que o açúcar alimenta o câncer: se as células cancerígenas precisam de muita glicose, cortar o açúcar da nossa dieta deve ajudar a impedir o crescimento do câncer e até mesmo impedir que ele se desenvolva em primeiro lugar. Infelizmente, não é tão simples assim. Todas as nossas células saudáveis também precisam de glicose, e não há como dizer aos nossos corpos para permitir que as células saudáveis tenham a glicose de que necessitam, mas não para as células cancerígenas.
E seguir dietas severamente restritas com quantidades muito baixas de carboidratos pode prejudicar a saúde a longo prazo, eliminando alimentos que são boas fontes de fibras e vitaminas.
Isto é particularmente importante para pacientes com câncer, porque alguns tratamentos podem resultar em perda de peso e colocar o corpo sob muito estresse. Portanto, a má nutrição das dietas restritivas também pode dificultar a recuperação ou até mesmo ameaçar a vida.
Um final “pegajoso” para pesquisa sobre açúcar?
Embora não haja evidência de que o corte de carboidratos da nossa dieta ajude a tratar o câncer, pesquisas importantes mostraram que entender as formas anormais pelas quais as células cancerosas produzem energia pode levar a novos tratamentos.
Nos anos 50, um cientista chamado Otto Warburg notou que as células cancerígenas usam um processo químico diferente das células normais para transformar glicose em energia.
Células saudáveis usam uma série de reações químicas em pequenas baterias celulares chamadas mitocôndrias. O Efeito Warburg, como foi apelidado após a descoberta de Otto, descreve como as células cancerígenas contornam suas "baterias" para gerar energia mais rapidamente para atender à demanda.
Esse atalho para gerar energia pode ser uma fraqueza para alguns tipos de câncer, o que dá aos pesquisadores uma vantagem para o desenvolvimento de novos tratamentos.
Em primeiro lugar, ele abre o potencial para o desenvolvimento de drogas que desligam os processos de produção de energia das células cancerígenas, mas não impedem que as células saudáveis produzam energia. E os pesquisadores estão testando drogas que funcionam dessa maneira.
Em segundo lugar, os processos anormais nas células cancerosas também podem deixá-los menos aptos a se adaptar diante da falta de outros nutrientes, como os aminoácidos. Essas vulnerabilidades potenciais também podem levar a tratamentos.
Mas essas abordagens ainda são experimentais, e ainda não sabemos se os tratamentos que prejudicam as células cancerosas são seguros ou se funcionam.
Certamente não é motivo para os pacientes com câncer tentarem fazer isso sozinhos restringindo sua dieta durante o tratamento - e voltando ao nosso ponto anterior, pode ser perigoso fazê-lo.
Se o açúcar não causar câncer, por que se preocupar com isso?
Cortar o açúcar não ajuda a tratar o câncer, e o açúcar não causa câncer diretamente. Por que, então, encorajamos as pessoas a reduzirem os alimentos açucarados em nosso aconselhamento sobre dieta?
Isso porque existe uma ligação indireta entre o risco de câncer e o açúcar. Comer muito açúcar com o tempo pode fazer com que você ganhe peso, e evidências científicas robustas mostram que o excesso de peso ou obesidade aumenta o risco de 13 tipos diferentes de câncer. De fato, a obesidade é a maior causa evitável de câncer após o cigarro, sobre a qual já escrevemos muitas vezes.
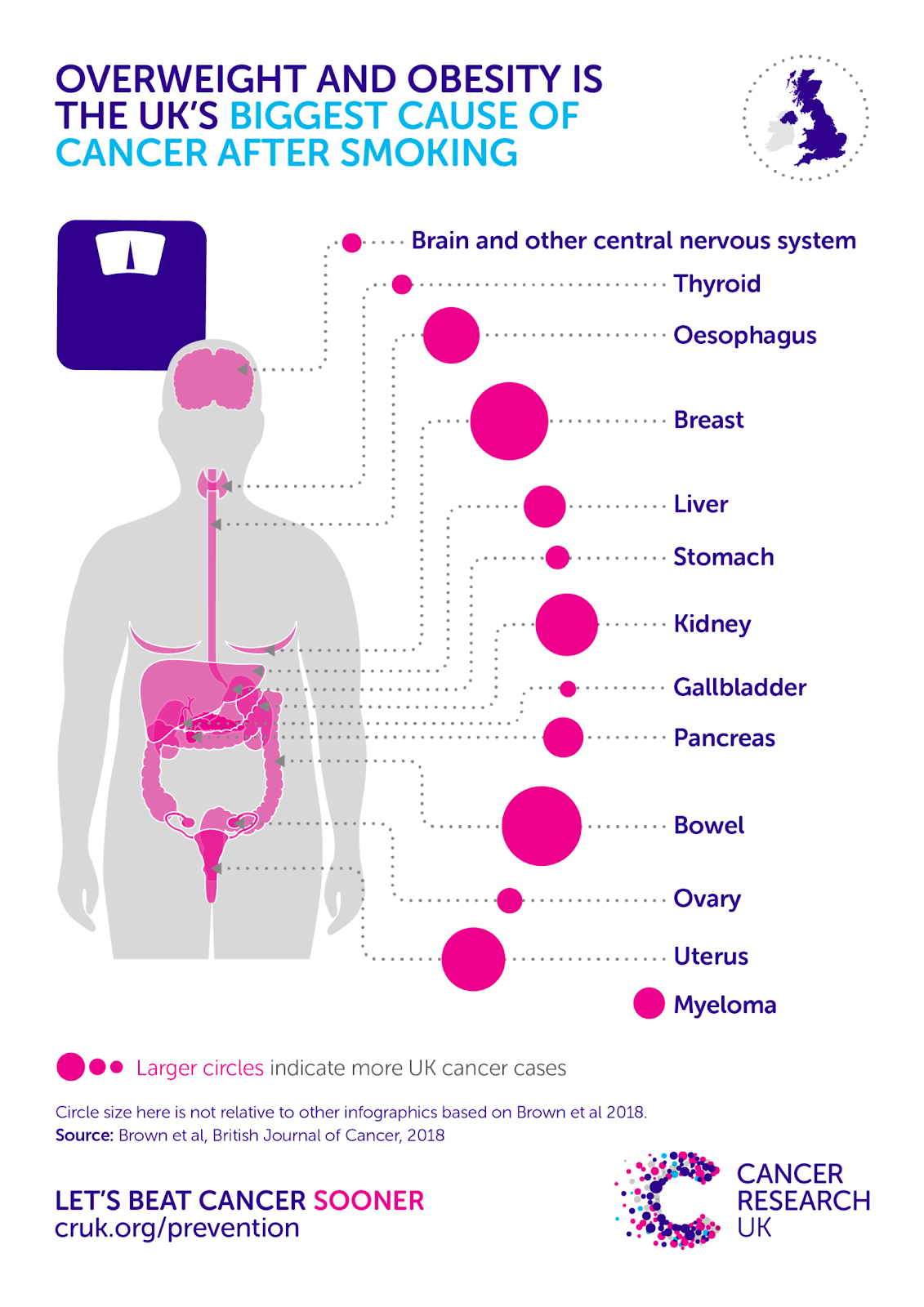
É o açúcar que mais nos preocupa quando se trata de ganho de peso, e não de açúcar que é encontrado naturalmente em alimentos como frutas e leite ou alimentos ricos em amido, como grãos integrais e legumes (que as pessoas deveriam comer mais *).
Como posso reduzir o açúcar adicionado?
Uma das maneiras mais fáceis de reduzir o seu açúcar adicionado é reduzir as bebidas açucaradas, que são a maior fonte de açúcar na dieta do Reino Unido.
Algumas bebidas açucaradas, como refrigerantes e bebidas energéticas, podem ter mais do que a quantidade máxima diária recomendada de açúcar adicionado em apenas uma porção. E enquanto essas calorias extras promovem ganho de peso, elas não oferecem nenhum outro benefício nutricional.
“Múltiplas dicas nos empurram como clientes para empilhar junk food em nossas cestas de compras, mesmo se não estivéssemos planejando” - Professora Linda Bauld, Cancer Research UK
Outros alimentos obviamente açucarados, como doces, chocolate, bolos e biscoitos são todos mantidos como guloseimas também. Mas alguns alimentos que escondem grandes quantidades de açúcar adicionado podem surpreendê-lo. Alguns cereais matinais, refeições prontas (incluindo os "saudáveis"), molhos para massas e iogurtes podem ter chocantes quantidades de açúcar adicionados a eles. Ler os rótulos de informações nutricionais e verificar a lista de ingredientes pode ajudá-lo a escolher opções de menor teor de açúcar.
Embora haja etapas que você e sua família possam tomar para reduzir o consumo de açúcar, fazer essas mudanças pode ser mais fácil de dizer do que fazer. E é aqui que os governos precisam ajudar.
"Múltiplas pistas nos empurram como clientes para empilhar junk food em nossas cestas de compras, mesmo se não estivéssemos planejando", diz Linda Bauld, nossa campeã de prevenção do câncer baseada na Universidade de Edimburgo. "É por isso que queremos que o governo ajude a criar um ambiente alimentar melhor, onde a escolha saudável é a escolha mais fácil para todos."
Estamos muito satisfeitos que o imposto sobre o açúcar, que entrou em vigor em abril de 2018, já tenha tido um impacto, com algumas das maiores empresas de bebidas mudando suas receitas para reduzir o açúcar. O imposto poderia evitar milhões de casos de obesidade e cânceres associados à obesidade no futuro, reduzindo a quantidade de açúcar que o país consome em refrigerantes.
Mas o governo não fez muito progresso em seu plano de reduzir a quantidade de açúcar nos tipos de alimentos que são muito populares entre as crianças. Um ano depois do início do programa, a indústria não conseguiu cumprir as metas voluntárias estabelecidas pelo governo, mostrando que uma abordagem voluntária simplesmente não é tão eficaz.
Sem terminações doces
A história sobre o açúcar e o câncer é complicada.
Por um lado, o açúcar em si não causa câncer, e não há nenhuma maneira (no momento) de células cancerígenas de glicose serem colocadas em estado de fome, sem prejudicar as células saudáveis também.
Também não há evidências de que a adoção de uma dieta com baixo teor de carboidratos diminua o risco de câncer ou ajude como tratamento. E para os pacientes, obter uma nutrição adequada é importante para ajudar seus corpos a lidar com o tratamento.
Mas estamos preocupados com a quantidade de açúcar adicionado que as pessoas estão consumindo porque está promovendo o ganho de peso. E estar acima do peso ou obeso aumenta o risco de pelo menos 13 tipos de câncer.
Assim, a mensagem de levar para casa é que, embora banir o açúcar não pare o câncer, todos nós podemos reduzir nosso risco de contrair câncer fazendo escolhas saudáveis e reduzir a quantidade de açúcar adicionado em nossas dietas é uma boa maneira de ajudar a manter o câncer. um peso corporal saudável.
* Enquanto alimentos como frutas, leite e alimentos ricos em amido são ricos em carboidratos, eles têm outros benefícios nutricionais importantes. Todos devemos comer mais frutas, verduras, legumes e leguminosas integrais, pois esses alimentos nutritivos também são ricos em fibras - isso não apenas ajuda seu corpo a digerir o açúcar natural mais lentamente (o que ajuda a manter um peso saudável), como também reduz o risco de câncer de intestino.
Adicionais
Observamos que posições moderadas sobre o tema são encontráveis na internet
Observamos que posições moderadas sobre o tema são encontráveis na internet
Destaque-se:
“A dieta cetogênica tem sido estudada como um tratamento adicional contra o câncer que, aliada à quimioterapia e radioterapia, pode ajudar a reduzir a progressão do tumor. Ela foi difundida no Brasil pelo médico e nutrólogo Lair Ribeiro, mas ainda são poucos os dados e estudos que confirmam a eficácia dessa dieta contra o câncer.
[...]
Ao fazer uma dieta cetogênica, o nível de glicose, que é o açúcar no sangue, fica bastante reduzido, e esse é único combustível que as células cancerosas conseguem processar para crescer e se multiplicar. Assim, é como se a dieta fizesse as células ficarem sem comida e com isso ajudasse a controlar o avanço da doença.
Além disso, o baixo teor de carboidratos também podem levar a níveis mais baixos de circulação dos hormônios insulina e IGF-1, podendo fazer com que as células cancerosas tenham menos sinais para crescer e dividir.
Por outro lado, as células saudáveis do corpo conseguem utilizar ácidos graxos e corpos cetônicos como fontes de energia, nutrientes que vêm da gordura da alimentação e das reservas de gordura do corpo."
[...]
A dieta cetogênica só deve ser feita em pacientes com câncer após consentimento do médico e com acompanhamento do nutricionista, sendo necessário observar o aparecimento de efeitos colaterais como tontura e fraqueza, especialmente nos primeiros dias.
Também é importante lembrar que os estudos relacionados à dieta cetogênica e câncer ainda não são conclusivos e que essa dieta não é adequada em todos os casos de câncer. Além disso, ela não substitui os tratamentos convencionais com medicamentos, quimioterapia, radioterapia ou hormonioterapia.”
O papel da restrição do carboidrato na dieta para a prevenção e o tratamento do câncer - Daniela Pessoa - 24 nov 2016 - vejario.abril.com.br
Nos nossos arquivos; [ Daniela Pessoa - Papel restrição carboidrato na dieta para prevenção e tratamento câncer ]
Destaque-se:
“A restrição de carboidratos mimetiza o estado metabólico de restrição calórica ou do jejum. Os efeitos benéficos da restrição calórica e do jejum sobre o risco de câncer e sua progressão estão bem estabelecidos. A restrição de carboidratos oferece, portanto, a possibilidade de alcançar os mesmos mecanismos subjacentes sem os efeitos secundários da fome e da perda de peso.
Existe, portanto, um papel importante da restrição do açúcar na dieta no combate ao câncer. Por isso, vários ensaios clínicos já utilizam a dieta restrita em carboidratos como uma opção terapêutica de suporte em oncologia.”
Infográfico útil
Rainer J Klement, Ulrike Kämmerer; Is There a Role for Carbohydrate Restriction in the Treatment and Prevention of Cancer?; Nutr Metab. 2011;8(75) - www.ncbi.nlm.nih.gov
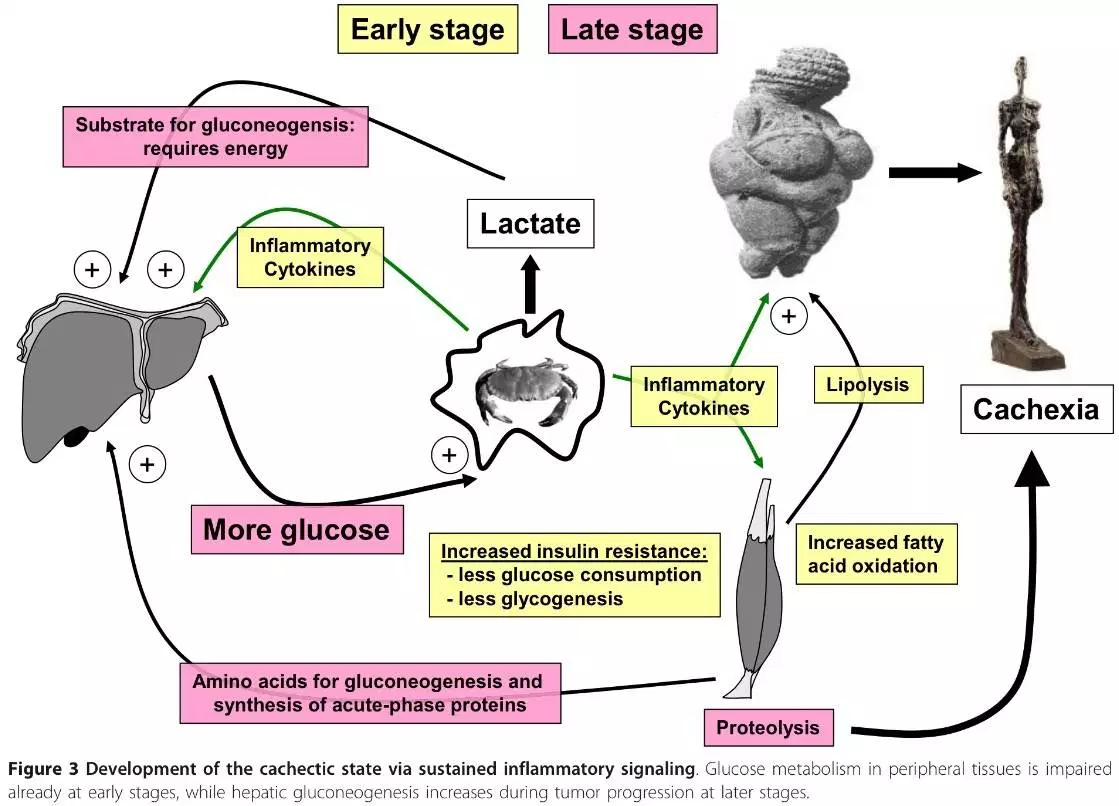
As conclusões deste artigo:
Resumimos nossos principais achados da literatura sobre o papel da restrição de CHO na dieta no desenvolvimento e no resultado do câncer.
(i) A maioria, se não todas, as células tumorais têm uma alta demanda de glicose em comparação com as células benignas do mesmo tecido e conduzem a glicólise mesmo na presença de oxigênio (o efeito Warburg). Além disso, muitas células cancerosas expressam receptores de insulina (IRs) e apresentam hiperativação da via do IGF1R-IR. Existe evidência de que os níveis cronicamente elevados de glicose, insulina e IGF1 no sangue facilitam a tumorigênese e pioram o desfecho em pacientes com câncer.
(ii) O envolvimento do eixo glicose-insulina também pode explicar a associação da síndrome metabólica com o aumento do risco de vários cânceres. Já foi demonstrado que a restrição de CHO (carboidratos) exerce efeitos favoráveis em pacientes com a síndrome metabólica. Estudos epidemiológicos e antropológicos indicam que a restrição de CHOs na dieta pode ser benéfica na redução do risco de câncer.
(iii) Muitos pacientes com câncer, em particular aqueles com estágios avançados da doença, apresentam metabolismo de todo o corpo alterado marcado por níveis plasmáticos aumentados de moléculas inflamatórias, síntese de glicogênio prejudicada, aumento da proteólise e aumento da utilização de gordura no tecido muscular, aumento da lipólise na gordura adiposa tecido e aumento da gliconeogênese pelo fígado. As dietas CHO com alto teor de gordura e baixo objetivo são responsáveis por essas alterações metabólicas. Estudos conduzidos até agora mostraram que tais dietas são seguras e provavelmente benéficas, em particular para pacientes com câncer em estágio avançado.
(iv) a restrição de CHO mimetiza o estado metabólico da restrição calórica ou - no caso de KDs (ketogenic diet, dieta cetogênica) - o jejum. Os efeitos benéficos da restrição calórica e do jejum no risco e progressão do câncer estão bem estabelecidos. A restrição CHO abre, assim, a possibilidade de visar os mesmos mecanismos subjacentes, sem os efeitos colaterais da fome e perda de peso.
(v) Alguns estudos laboratoriais indicam um potencial antitumoral direto de corpos cetônicos. Durante os últimos anos, uma infinidade de estudos com camundongos realmente comprovaram os efeitos antitumorais dos KDs para vários tipos de tumores, e alguns relatos de casos e estudos pré-clínicos também obtiveram resultados promissores em pacientes com câncer. Vários ensaios clínicos registrados vão investigar o caso de uma KD como uma opção terapêutica de suporte em oncologia.
Para os não muito familiarizados com o papel dos carboidratos na geração de energia para atletas (caso elevado do que ocorre em qualquer pessoa), recomendo meu texto:
Carboidratos e geração de energia - Uma abordagem didática para praticantes de musculação - Scientia